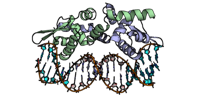
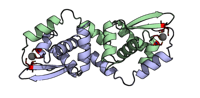
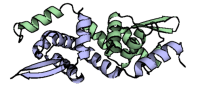
3 passes of 1440 raw diffraction images modeled by PDB ID 9B7C.
Data DOI: 10.15785/SBGRID/1276 | PDB ID 9B7C: RCSB PDBe | Publication DOI: 10.1126/sciadv.adj2921 | Published: 7 Jul 2026
Hekstra Laboratory, Harvard University
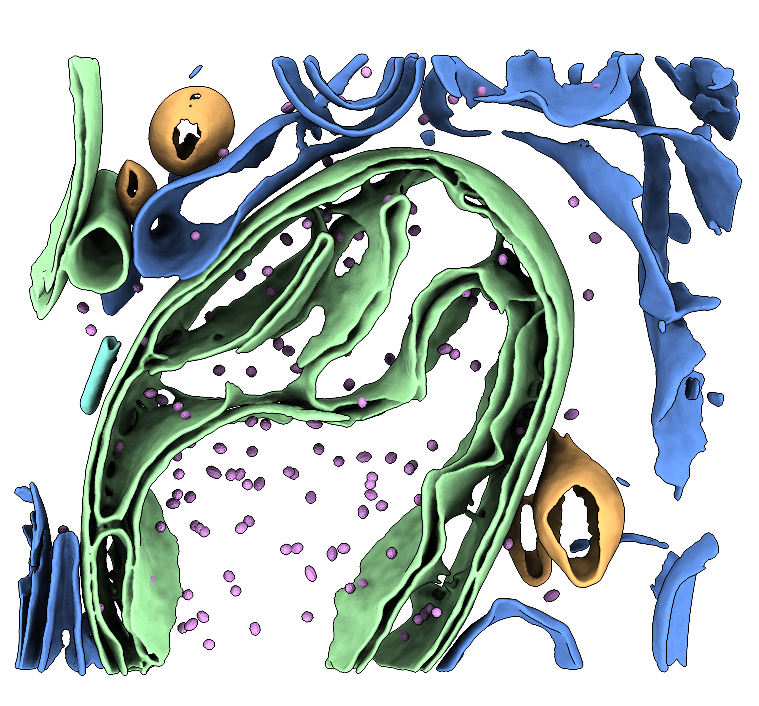
Two raw tilt-series (in MRC format) of FIB-milled G-TPP-treated HeLa cells that have been milled. The dataset contains mitochondria and cytoplasmic ribosomes as well as mHSP60-mHSP10 chaperonin "football" complexes. MRC files and MDOC files are provided. See the Processing Bundle below for all the scripts necessary to reconstruct the data to low-resolution following our tutorial. This tutorial involves template matching, subtomogram averaging, membrane segmentation, and particle place-back. See the processing instruction document for full details. Storage required: 4.4 G (dataset size) and 50G (storage for processing files when the tutorial has been completed).
Data DOI: 10.15785/SBGRID/1273 | Publication DOI: 10.1101/2025.10.06.680693 | Published: 10 Jul 2026
Ruben Fernandez-Busnadiego, University of Göttingen
X-ray diffraction images for Glutarate hydroxylase (GlaH) from Escherichia coli
Data DOI: 10.15785/SBGRID/1271 | PDB ID 8CDF: RCSB PDBe | Published: 7 Jul 2026
Schubert Laboratory, University of Pretoria
X-Ray Diffraction data from M. hassiacum GgH (Q434F mutant), source of 6G3N structure
Data DOI: 10.15785/SBGRID/1269 | PDB ID 6G3N: RCSB PDBe | Published: 8 May 2026
Pereira Laboratory, IBMC/i3S, Universidade do Porto
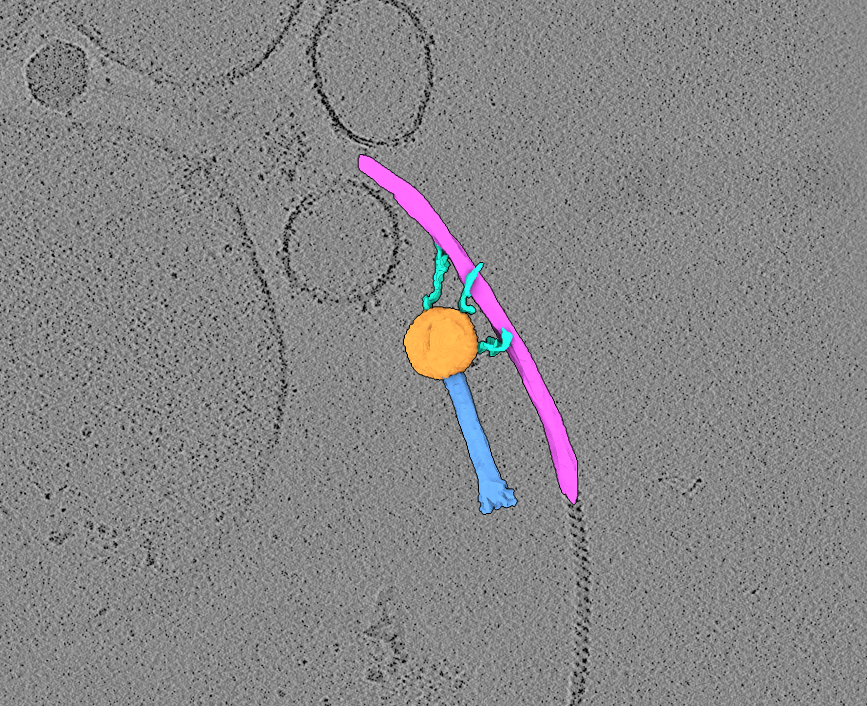
Two raw tilt-series (in MRC format) of highly-purified jumbo bacteriophage in association with bacterial flagella. MDOC files are also provided.
Data DOI: 10.15785/SBGRID/1263 | Publication DOI: doi.org/10.1038/s42003-025-09319-7 | Published: 21 Apr 2026
Briegel Laboratory, Institut Pasteur
Crystal Structure of the Listeria monocytogenes CadC in complex with DNA
Data DOI: 10.15785/SBGRID/1261 | PDB ID 9T6T: RCSB PDBe | Published: 28 Apr 2026
Cabanes Laboratory, i3S
Crystal Structure of the Listeria monocytogenes CadC with Cadmium
Data DOI: 10.15785/SBGRID/1260 | PDB ID 9T6S: RCSB PDBe | Published: 28 Apr 2026
Cabanes Laboratory, i3S
Crystal Structure of the Listeria monocytogenes CadC
Data DOI: 10.15785/SBGRID/1259 | PDB ID 9T6R: RCSB PDBe | Published: 28 Apr 2026
Cabanes Laboratory, i3S
Native dataset. The crystal was macroscopically twinned.
Data DOI: 10.15785/SBGRID/1256 | PDB ID 8EVD: RCSB PDBe | Published: 10 Apr 2026
Madden Laboratory, Dartmouth Geisel School of Medicine
Tilt-series of milled Saccharomyces cerevisiae cells depicting mitochondria, nuclear envelopes, lipid bodies, smooth endoplasmic reticulum and other intracellular components.
Data DOI: 10.15785/SBGRID/1255 | Published: 21 Apr 2026
Raunser Laboratory, Max Planck Institute of Molecular Physiology
Native dataset
Data DOI: 10.15785/SBGRID/1241 | PDB ID 9Z7O: RCSB PDBe | Published: 26 May 2026
Corbett Laboratory, University of California, San Diego
Native dataset
Data DOI: 10.15785/SBGRID/1240 | PDB ID 9Z73: RCSB PDBe | Published: 26 May 2026
Corbett Laboratory, University of California, San Diego
Native dataset
Data DOI: 10.15785/SBGRID/1239 | PDB ID 9Z72: RCSB PDBe | Published: 26 May 2026
Corbett Laboratory, University of California, San Diego
Native dataset
Data DOI: 10.15785/SBGRID/1238 | PDB ID 9Z71: RCSB PDBe | Published: 26 May 2026
Corbett Laboratory, University of California, San Diego
Data collected at Vanadium peak wavelength
Data DOI: 10.15785/SBGRID/1235 | Published: 2 Dec 2025
Drennan Laboratory, Massachusetts Institute of Technology
Native dataset
Data DOI: 10.15785/SBGRID/1234 | PDB ID 9PCN: RCSB PDBe | Published: 2 Dec 2025
Drennan Laboratory, Massachusetts Institute of Technology
Native dataset
Data DOI: 10.15785/SBGRID/1233 | PDB ID 9PCM: RCSB PDBe | Published: 2 Dec 2025
Drennan Laboratory, Massachusetts Institute of Technology
Native dataset
Data DOI: 10.15785/SBGRID/1232 | PDB ID 9PCO: RCSB PDBe | Published: 2 Dec 2025
Drennan Laboratory, Massachusetts Institute of Technology
Native dataset
Data DOI: 10.15785/SBGRID/1231 | PDB ID 9PCL: RCSB PDBe | Published: 2 Dec 2025
Drennan Laboratory, Massachusetts Institute of Technology
Native dataset
Data DOI: 10.15785/SBGRID/1230 | PDB ID 9PC5: RCSB PDBe | Published: 2 Dec 2025
Drennan Laboratory, Massachusetts Institute of Technology